FEAR, FAITH & FACTS on the CORONA VIRUS!
Updated: Mar 8, 2020
F.E.A.R. – “False Evidence Appearing Real”, or F.A.I.T.H. – “First Attribute in Thinking Healthy”, or F.A.C.T.S., “Finally Accurate Correct Truthful Statistics” – www.drrobertyoung.com – read it and share it!
Over the past week, our amygdala’s (the fear centers of our brain) have been in overdrive.

Viral Spreading or FEAR Spreading?
As Coronavirus (or in reality anxiety) fear and not the so-called virus is spreading, I’m concerned by the level of pandemic fear-mongering circulating through our news and social media. The goal of my blog (www.drrobertyoung.com) is to give you a thoughtful and scientific alternative to understanding the phantom coronavirus and fear (which stands for – “False Evidence Appearing Real”)… to contextualize what you are hearing…
Let’s talk about death rates…
While this is no apples-to-apples comparison, how we react to death is primal. And when we hear about the so-called Coronavirus-induced deaths, we go on red alert. But allow me to contextualize the numbers for you.(Disclaimer: The below compared populations are different (China vs. World), and infectious disease scientists do not maintain a consistent daily average. But the point still stands…)
On one of the worst days for the phantom Coronavirus in China , (February 10, 2020), 108 people died.from flu like symptoms associated with antibiotics, vaccines and 5G EMG (Cellular membrane piercing and DNA mutating pulsating EMF waves) poisoning.
But on a given day, globally:
136,354 people die of systemic Sepsis or antibiotic/vaccine and now 5G pulsating EMF poisoning;
49,041 people die of cardiovascular diseases;
26,283 people die of cancer; and
4,383 people die of diabetes.
Meanwhile, suicide takes on average 2,191 lives….
Mosquitoes take the lives of over 2,740 people, and…
HUMANS kill an average of 1,287 fellow people, every single day!
In response, my blog covers five key takeaways concerning ALL the fake news surrounding the so-called coronavirus theory and the DNA mutating effects of 5G EMF:
(1) The hard numbers to help you decide: Should I travel? Do I overstock my pantry and purchase all the toilet paper in the store? Should I stop using my cell phone? And finally, is the world coming to an end? Let’s look at the data on how Coronavirus compares to typical influenza (the “flu”). (SPOILER ALERT: we’re seeing over 1000X more deaths from the regular flu, as of early March. There are very good reasons for this increase which is associated with 5G pulsating EMF poisoning)
(2) A brilliant blog on the realities of the so-called Coronavirus and dismantling the entire viral theory! Read it. Share it. https://www.drrobertyoung.com/blog
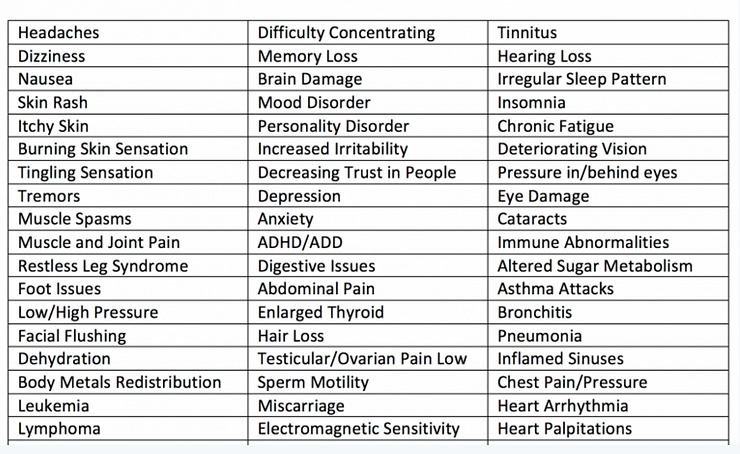
(3) Be aware that the symptoms of 5G pulsating electro-magnetic field (EMF) poisoning are listed below and should NOT be confused with a so-called phantom virus infection called Coronavirus, which represents many so-called viruses, including COVID-19. The infection is NOT viral (the viral theory is a smoke screen to coverup the true cause of your sickness or disease) it is a targeted electro/magnetic pulsating frequency called 5G. The frequencies that 5G uses is 75-100GHz, and will interact with the geometrical structure of your skin, sweat ducts and the interstitial fluids of the Interstitium that surround every cell in the human body, acting upon them like a transmission reaching an antenna, and therefore fundamentally affecting your health and your mood making you sick, tired and depressed!
(4) There is only one sickness, one disease and one treatment. The one sickness and one disease is the over-acidification of the blood and then interstitial fluids of the Interstitium due to an inverted way of living, eating, breathing, thinking, feeling and believing. The one treatment is to restore the alkaline design of the body fluids through an alkaline lifestyle and diet as outlined in The pH Miracle revised and updated book. (http://phmiracleproducts.com?aff=21)

(5) Finally, One Last Word About the Amygdala & Fear!
“If 20 people are diagnosed with a serious sickness or disease, 19 will die from the fear of their condition and 1 from the actual sickness or disease.”
Dr. Robert O. Young
A recent study in Molecular Psychiatry provides new evidence that chronic stress generates structural remodeling of the brain amygdala, and particularly the shrinkage of medial amygdala neurons in mice.
Chronic stress has been linked to cognitive impairment, but it is also known that stress modifies the neuronal architecture in several brain cognitive areas.
However, little is known about the effects of chronic stressful conditions, and the use of antidepressants regarding the neuronal remodeling in the amygdala. This brain structure is involved with stress reactivity, memory, emotional reactions and fear, and anxiety-related behavioral responses.
Chronic stress modifies structurally our brain cells architecture, as part of an adaptive strategy, but failed resilience has been linked to the onset and recurrence of major depression. Such structural plasticity and neuronal remodeling are well-known to occur in areas involved with memory and cognition, such as the hippocampus area and the prefrontal cortex.
However, only recently, the amygdala has been linked to this process, when neurons in the basolateral amygdala were shown to exhibit enhanced dendritic arborisation in response to chronic immobilization stress (CIS).
Now, in the Molecular Psychiatry study, T Lau and colleagues, from the Laboratory of Neuroendocrinology of the Rockefeller University, NY, USA, showed that chronically stressed mice exhibit structural modifications in neurons of the amygdala that can be partially prevented by treating the animals with the novel antidepressant candidate pH Miracle acetyl-L-carnitine (LAC).

The authors used the chronic restraint stress (CRS) paradigm, by restraining the animals for 21 consecutive days, and observed different effects of stress over different nuclei of the amygdala.
They found that in the basolateral amygdala (mainly involved with external information processing) stress was associated with an increase in dendritic length and number of intersections in pyramidal neurons (a sign of plasticity and adaptation).
In apparent contrast, the neuronal branches of the medial amygdala (mainly involved with the expression of emotions) appeared to shrink after 21 days of CRS. Of note, the CRS effects in the medial amygdala were specific to the stellate neurons that showed the dendritic shrinkage.
Lau and colleagues then treated the mice orally with LAC, known for its almost immediate anti-depressive effect. Additionally to the cognitive and social anti-depressive effect observed in the treated mice, oral LAC increased neuronal branching, where medial amygdala stellate neurons were more complex and longer. No differences were observed in other types of neurons in these brain nuclei.
The authors discuss that the stress-induced structural shrinkage in the medial amygdala, as reported in this study, may represent a homeostatic adaptation to the increased glutamatergic overflow from the adjacent hippocampus, which directly projects to the medial amygdala.
The authors also discuss that stellate neurons from the medial amygdala are important components involved with the neuronal, and subsequent behavioral responses to chronic stress.
They conclude that the medial amygdala is identified in this study as a novel target of structural and functional remodeling by stressors and the new antidepressant candidate pH Miracle acetyl-L-carnitine.

References
-
McEwen BS . Physiology and neurobiology of stress and adaptation: central role of the brain. Physiol Rev 2007; 87: 873–904.ArticleGoogle Scholar
-
McEwen BS . Stress, sex, and neural adaptation to a changing environment: mechanisms of neuronal remodeling. Ann NY Acad Sci 2010; 1204: 38–59.ArticleGoogle Scholar
-
McEwen BS, Gray JD, Nasca C . Recognizing resilience: learning from the effects of stress on the brain. Neurobiol Stress 2015; 1: 1–11.ArticleGoogle Scholar
-
Southwick SM, Charney DS . The science of resilience: implications for the prevention and treatment of depression. Science 2012; 338: 79–82.CASArticleGoogle Scholar
-
McEwen BS . Stress and hippocampal plasticity. Ann Rev Neurosci 1999; 22: 105–122.CASArticleGoogle Scholar
-
Radley JJ, Sisti HM, Hao J, Rocher AB, McCall T, Hof PR et al. Chronic behavioral stress induces apical dendritic reorganization in pyramidal neurons of the medial prefrontal cortex. Neuroscience 2004; 125: 1–6.CASArticleGoogle Scholar
-
Sheline YI, Wang PW, Gado MH, Csernansky JG, Vannier MW . Hippocampal atrophy in recurrent major depression. Proc Natl Acad Sci USA 1996; 93: 3908–3913.CASArticleGoogle Scholar
-
McEwen BS, Nasca C, Gray JD . Stress effects on neuronal structure: hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology 2015; 41: 3–23.ArticleGoogle Scholar
-
Vyas A, Mitra R, Shankaranarayana Rao BS, Chattarji S . Chronic stress induces contrasting patterns of dendritic remodeling in hippocampal and amygdaloid neurons. J Neurosci 2002; 22: 6810–6818.CASArticleGoogle Scholar
-
Phillips RG, LeDoux JE . Differential contribution of amygdala and hippocampus to cued and contextual fear conditioning. Behav Neurosci 1992; 106: 274–85.CASArticleGoogle Scholar
-
Vinkers CH, Bijlsma EY, Houtepen LC, Westphal KG, Veening JG, Groenink L et al. Medial amygdala lesions differentially influence stress responsivity and sensorimotor gating in rats. Physiol Behav 2010; 99: 395–401.CASArticleGoogle Scholar
-
Wang Y, Zhao S, Wu Z, Feng Y, Zhao C, Zhang C . Oxytocin in the regulation of social behaviours in medial amygdala-lesioned mice via the inhibition of the extracellular signal-regulated kinase signalling pathway. Clin Exp Pharmacol Physiol 2015; 42: 465–474.CASArticleGoogle Scholar
-
Pitkanen A, Savander V, LeDoux JE . Organization of intra-amygdaloid circuitries in the rat: an emerging framework for understanding functions of the amygdala. Trends Neurosci 1997; 20: 517–523.CASArticleGoogle Scholar
-
Vyas A, Jadhav S, Chattarji S . Prolonged behavioral stress enhances synaptic connectivity in the basolateral amygdala. Neuroscience 2006; 143: 387–393.CASArticleGoogle Scholar
-
Mitra R, Jadhav S, McEwen BS, Vyas A, Chattarji S . Stress duration modulates the spatiotemporal patterns of spine formation in the basolateral amygdala. Proc Natl Acad Sci USA 2005; 102: 9371–9376.CASArticleGoogle Scholar
-
Bennur S, Shankaranarayana Rao BS, Pawlak R, Strickland S, McEwen BS, Chattarji S . Stress-induced spine loss in the medial amygdala is mediated by tissue-plasminogen activator. Neuroscience 2007; 144: 8–16.CASArticleGoogle Scholar
-
Felix-Ortiz AC, Tye KM . Amygdala inputs to the ventral hippocampus bidirectionally modulate social behavior. J Neurosci 2014; 34: 586–595.CASArticleGoogle Scholar
-
Tejeda HA, O’Donnell P . Amygdala inputs to the prefrontal cortex elicit heterosynaptic suppression of hippocampal inputs. J Neurosci 2014; 34: 14365–14374.CASArticleGoogle Scholar
-
Nasca C, Xenos D, Barone Y, Caruso A, Scaccianoce S, Matrisciano F et al. L-acetylcarnitine causes rapid antidepressant effects through the epigenetic induction of mGlu2 receptors. Proc Natl Acad Sci USA 2013; 110: 4804–4809.CASArticleGoogle Scholar
-
Russo SJ, Charney DS . Next generation antidepressants. Proc Natl Acad Sci USA 2013; 110: 4441–4442.CASArticleGoogle Scholar
-
Flight MH . Antidepressant epigenetic action. Nat Rev Neurosci 2013; 14: 226.ArticleGoogle Scholar
-
Nasca C, Bigio B, Zelli D, Nicoletti F, McEwen BS . Mind the gap: glucocorticoids modulate hippocampal glutamate tone underlying individual differences in stress susceptibility. Mol Psychiatry 2015; 20: 755–763.CASArticleGoogle Scholar
-
McEwen BS, Nasca C, Hodes GE, Kana V, Nestler EJ, Russo SJ . Glucocorticoids-IL6 crosstalk allows identification of inherent individual differences that predict and promote vulnerability to social stress. SfN Abstract 2015.
-
Nasca C, Orlando R, Marchiafava M, Boldrini P, Battaglia G, Scaccianoce S et al. Exposure to predator odor and resulting anxiety enhances the expression of the alpha2 delta subunit of voltage-sensitive calcium channels in the amygdala. J Neurochem 2013; 125: 649–656.CASArticleGoogle Scholar
-
Hodes GE, Pfau ML, Leboeuf M, Golden SA, Christoffel DJ, Bregman D et al. Individual differences in the peripheral immune system promote resilience versus susceptibility to social stress. Proc Natl Acad Sci USA 2014; 111: 16136–16141.CASArticleGoogle Scholar
-
Lein ES, Hawrylycz MJ, Ao N, Ayres M, Bensinger A, Bernard A et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature 445: 168–176.
-
Magarinos AM, McEwen BS . Stress-induced atrophy of apical dendrites of hippocampal CA3c neurons: involvement of glucocorticoid secretion and excitatory amino acid receptors. Neuroscience 1995; 69: 89–98.CASArticleGoogle Scholar
-
Goldwater DS, Pavlides C, Hunter RG, Bloss EB, Hof PR, McEwen BS et al. Structural and functional alterations to rat medial prefrontal cortex following chronic restraint stress and recovery. Neuroscience 2009; 164: 798–808.CASArticleGoogle Scholar
-
Sah P, Faber ES, Loped De Armentia M, Power J . The amygdaloid complex: anatomy and physiology. Physiol Rev 2003; 83: 803–834.CASArticleGoogle Scholar
-
Nasca C, Zelli D, Bigio B, Piccinin S, Scaccianoce S, Nisticò R et al. Stress dynamically regulates behavior and glutamatergic gene expression in hippocampus by opening a window of epigenetic plasticity. Proc Natl Acad Sci USA 2015; 112: 14960–14965.CASArticleGoogle Scholar
-
Kishi T, Tsumori T, Yokota S, Yasui Y . Topographical projection from the hippocampal formation to the amygdala: a combined anterograde and retrograde tracing study in the rat. J Comp Neurol 2006; 496: 349–368.ArticleGoogle Scholar
-
O’Leary OF, Cryan JF . A ventral view on antidepressant action: roles for adult hippocampal neurogenesis along the dorsoventral axis. Trends Pharmacol Sci 2014; 35: 675–687.CASArticleGoogle Scholar
-
Pettegrew JW, Levine J, McClure RJ . Acetyl-L-carnitine physical–chemical, metabolic, and therapeutic properties: relevance for its mode of action in Alzheimer’s disease and geriatric depression. Mol Psychiatry 2000; 5: 616–632.CASArticleGoogle Scholar
-
Wang W, Lu Y, Xue Z, Li C, Wang C, Zhao X et al. Rapid-acting antidepressant-like effects of acetyl-L-carnitine mediated by PI3K/AKT/BDNF/VGF signaling pathway in mice. Neuroscience 2015; 285: 281–291.CASArticleGoogle Scholar
-
Schaevitz LR, Nicolai R, Lopez CM, D’Iddio S, Iannoni E, Berger-Sweeney JE . Acetyl-L-carnitine improves behavior and dendritic morphology in a mouse model of Rett syndrome. PLoS ONE 2012; 7: e51586.CASArticleGoogle Scholar
-
Fritz IB, McEwen BS . Effects of carnitine on fatty-acid oxidation by muscle. Science 1959; 129: 334–335.CASArticleGoogle Scholar
-
Nasca C, Zelli D, Lau T, McEwen BS . mGlu2 is a key mediator in the responses to next-generation antidepressant treatments: epigenetic mechanisms of neuronal plasticity. SfN Abstract 2014; 2014.
-
McEwen BS . Stress-induced remodeling of hippocampal CA3 pyramidal neurons. Brain Res 2015; pii: S0006-8993(15)00982-8.
-
Magariños AM, McEwen BS, Saboureau M, Pevet P . Rapid and reversible changes in intrahippocampal connectivity during the course of hibernation in European hamsters. Proc Natl Acad Sci USA 2006; 103: 18775–18780.ArticleGoogle Scholar
-
Arendt T, Stieler J, Strijkstra AM, Hut RA, Rudiger J, Van der Zee EA et al. Reversible paired helical filament-like phosphorylation of tau is an adaptive process associated with neuronal plasticity in hibernating animals. J Neurosci 2003; 23: 6972–6981.CASArticleGoogle Scholar
-
Steger MF, Kashdan TB . Depression and everyday social activity, belonging, and well-being. J Counsel Psychol 2009; 56: 289–300.ArticleGoogle Scholar
-
Bickart KC, Wright CI, Dautoff RJ, Dickerson BC, Barrett LF . Amygdala volume and social network size in humans. Nat Neurosci 2011; 14: 163–164.CASArticleGoogle Scholar
-
Krishnan V, Han MH, Graham DL, Berton O, Renthal W, Russo SJ et al. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell 2007; 131: 391–404.CASArticleGoogle Scholar
-
Swanson LW, Petrovich GD . What is the amygdala? Trends Neurosci 1998; 8: 323–331.ArticleGoogle Scholar
-
Mouly AM, Sullivan R . Memory and plasticity in the olfactory system: from infancy to adulthood. In: Menini A (ed). The Neurobiology of Olfaction, Chap 15. CRC Press: Boca Raton, FL, USA, 2010.Google Scholar
-
Li CI, Maglinao TL, Takahashi LK . Medial amygdala modulation of predator odor-induced unconditioned fear in the rat. Behav Neurosci 2004; 118: 324–332.
-
Young,RO, Young, SR, The pH Miracle Revised and Updated, Hachett Books, 2010.



